Author response: BNP facilitates NMB-encoded histaminergic itch via NPRC-NMBR crosstalk Article Swipe
 YOU?
·
· 2021
· Open Access
·
· DOI: https://doi.org/10.7554/elife.71689.sa2
· OA: W4206987834
YOU?
·
· 2021
· Open Access
·
· DOI: https://doi.org/10.7554/elife.71689.sa2
· OA: W4206987834
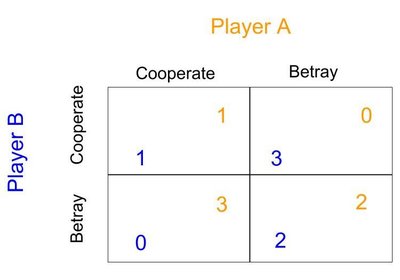
Article Figures and data Abstract Editor's evaluation eLife digest Introduction Results Discussion Materials and methods Data availability References Decision letter Author response Article and author information Metrics Abstract Histamine-dependent and -independent itch is conveyed by parallel peripheral neural pathways that express gastrin-releasing peptide (GRP) and neuromedin B (NMB), respectively, to the spinal cord of mice. B-type natriuretic peptide (BNP) has been proposed to transmit both types of itch via its receptor NPRA encoded by Npr1. However, BNP also binds to its cognate receptor, NPRC encoded by Npr3 with equal potency. Moreover, natriuretic peptides (NP) signal through the Gi-couped inhibitory cGMP pathway that is supposed to inhibit neuronal activity, raising the question of how BNP may transmit itch information. Here, we report that Npr3 expression in laminae I-II of the dorsal horn partially overlaps with NMB receptor (NMBR) that transmits histaminergic itch via Gq-couped PLCβ-Ca2+ signaling pathway. Functional studies indicate that NPRC is required for itch evoked by histamine but not chloroquine (CQ), a nonhistaminergic pruritogen. Importantly, BNP significantly facilitates scratching behaviors mediated by NMB, but not GRP. Consistently, BNP evoked Ca2+ responses in NMBR/NPRC HEK 293 cells and NMBR/NPRC dorsal horn neurons. These results reveal a previously unknown mechanism by which BNP facilitates NMB-encoded itch through a novel NPRC-NMBR cross-signaling in mice. Our studies uncover distinct modes of action for neuropeptides in transmission and modulation of itch in mice. Editor's evaluation The study by Meng et al., reveals how two distinct neuropeptide signals intersect to drive histaminergic itch. They find that the neuropeptides B-type natriuretic peptide (BNP) and neuromedin B (NMB) crosstalk in the spinal cord, whereby BNP facilitates itch behaviors driven by NMB via coupling between the receptors NPRC and NMBR. These results demonstrate a mechanism underlying spinal cord itch processing. https://doi.org/10.7554/eLife.71689.sa0 Decision letter eLife's review process eLife digest An itch is a common sensation that makes us want to scratch. Most short-term itches are caused by histamine, a chemical that is released by immune cells following an infection or in response to an allergic reaction. Chronic itching, on the other hand, is not usually triggered by histamine, and is typically the result of neurological or skin disorders, such as atopic dermatitis. The sensation of itching is generated by signals that travel from the skin to nerve cells in the spinal cord. Studies in mice have shown that the neuropeptides responsible for delivering these signals differ depending on whether or not the itch involves histamine: GRPs (short for gastrin-releasing proteins) convey histamine-independent itches, while NMBs (short for neuromedin B) convey histamine-dependent itches. It has been proposed that another neuropeptide called BNP (short for B-type natriuretic peptide) is able to transmit both types of itch signals to the spinal cord. But it remains unclear how this signaling molecule is able to do this. To investigate, Meng, Liu, Liu, Liu et al. carried out a combination of behavioral, molecular and pharmacological experiments in mice and nerve cells cultured in a laboratory. The experiments showed that BNP alone cannot transmit the sensation of itching, but it can boost itching signals that are triggered by histamine. It is widely believed that BNP activates a receptor protein called NPRA. However, Meng et al. found that the BNP actually binds to another protein which alters the function of the receptor activated by NMBs. These findings suggest that BNP modulates rather than initiates histamine-dependent itching by enhancing the interaction between NMBs and their receptor. Understanding how itch signals travel from the skin to neurons in the spinal cord is crucial for designing new treatments for chronic itching. The work by Meng et al. suggests that treatments targeting NPRA, which was thought to be a key itch receptor, may not be effective against chronic itching, and that other drug targets need to be explored. Introduction How itch and pain information is encoded and transmitted has been subjected to numerous studies for more than a century (Chen, 2021). There is increasing evidence indicating the pivotal roles of neuropeptides in the coding of itch information in primary sensory neurons (Chen, 2021). A pruritogenic stimulus activates skin, immune, and nerve cells, or an inflammatory response, which provokes the release of itch-specific neuropeptides from primary afferents to activate G-protein-coupled receptors (GPCRs) in the spinal cord (Chen, 2021; Wang and Kim, 2020). Notably, gastrin-releasing peptide (GRP) and neuromedin B (NMB), two mammalian neuropeptides, have been shown to encode nonhistaminergic itch and histaminergic itch, respectively (Akiyama et al., 2014; Barry et al., 2020; Sun and Chen, 2007; Wan et al., 2017; Zhao et al., 2014b). Moreover, murine GRP-GRPR signaling is important for the development of contact dermatitis-induced itch (Chen et al., 2020; Liu et al., 2020; Shiratori-Hayashi et al., 2015; Zhao et al., 2013). These findings are in accordance with human and animal studies showing that histaminergic and nonhistaminergic itch is transmitted through parallel primary afferent pathways (Chen, 2021; Johanek et al., 2007; Namer et al., 2008; Roberson et al., 2013; Wilson et al., 2011). B-type or brain natriuretic peptide (BNP), encoded by the gene Nppb, has been implicated in itch at discrete regions, including skin cells, sensory neurons, and spinal cord (Liu et al., 2020; Meng et al., 2018; Mishra and Hoon, 2013; Solinski et al., 2019). The natriuretic peptide (NP) family also consists of atrial (ANP) and C-type natriuretic peptides (CNP) (Potter et al., 2006). BNP binds to both NPRA and NPRC, encoded by Npr1 and Npr3, respectively, with equality affinity, but not NPRB, while ANP also binds NPRA directly, resulting in the elevation of the second message cyclic GMP concentration (Figure 1A; Potter et al., 2006). Although NPRC is considered to function as a clearance or silent receptor (Maack et al., 1987), it can also mediate guanylyl cyclase (GC) receptor-coupled Gαisignaling under certain physiological conditions (Anand-Srivastava, 2005). BNP-NPRA signaling was initially proposed as an itch-specific pathway responsible for transmitting both histamine- and CQ-evoked itch that acts upstream of GRP-GRPR signaling (Huang et al., 2018; Mishra and Hoon, 2013). However, that BNP transmits all types of itch is at odds with the fact that GRP is required only for nonhistaminergic itch. Further, genetic ablation of spinal Grp neurons fails to impact itch behaviors (Barry et al., 2020), indicating that spinal Grp neurons do not constitute a functional circuit for itch. Recent studies have shown that BNP-NPRA signaling is involved in histaminergic itch as well as chronic itch in mice which comprises the histaminergic component (Liu et al., 2020; Solinski et al., 2019). Given that BNP can bind both NPRA and NPRC, two cognate receptors for BNP (Figure 1A), the relationship between NPRA/NPRC and NMBR/GRPR and the role of NPRC in itch transmission remains undefined. Considering that the GC-cGMP signal transduction pathway mediated by BNP is inhibitory (Potter et al., 2006) and that BNP-NPRA/NPRC signaling may exert an inhibitory rather than excitatory function, analogous to GαI protein-coupled signaling, it is paradoxical that BNP would transmit rather than inhibiting itch information. Figure 1 with 2 supplements see all Download asset Open asset Expression of Npr1, 2,and 3 and other molecular markers in the spinal cord. (A) Diagram shows crosstalk between NPs and NP receptors. BNP can bind NPRA and NPRC. (B) BNP dose-dependently evoked scratching behaviors 60 min after i.t. injection. n = 6. *p < 0.05, **p < 0.01, one-way ANOVA followed by Tukey’s test. (C) Time-course of scratching behaviors induced by different doses of BNP shows a delayed onset of scratching responses. (D, F, H, J, L, N) Images of double RNAscope ISH showing that the overlapping expression of Npr1 (green) with Grpr (red) (D), Nmbr (F), of Npr3 (green) with Nmbr (red) (H), Npr3 (red) with Vglut2 (green) (J), Vgat (green) (L), or Npr1 (green) (N) in laminae I-II of the dorsal horn. Dashed white lines divide laminae I-II from III. White boxes are shown at higher magnification in the right panel. Arrows indicate double-positive neurons. E, G, I, K, M, O, Venn diagrams showing the overlap between Npr1 and Grpr (E), Nmbr (G), between Npr3 and Nmbr (I), Vglut2 (K), Vgat (M) or Npr1 (O). n = 10–15 sections from 3 mice. Scale bar, 20 µm in D – N. Figure 1—source data 1 BNP dose-dependently evoked scratching behaviors and showed a delayed onset of scratching responses. https://cdn.elifesciences.org/articles/71689/elife-71689-fig1-data1-v2.xlsx Download elife-71689-fig1-data1-v2.xlsx In the present study, we have examined these open questions using a combination of RNA-scope ISH, genetic knockout (KO) mice, spinal siRNA knockdown, cell ablation, calcium imaging, pharmacological and optogenetic approaches. We found that NPRC, rather than NPRA, is a major functional receptor for BNP in the spinal cord, and BNP facilitates histamine-evoked itch through NPRC-NMBR crosstalk. Importantly, our studies confirmed that BNP is an inhibitory neuropeptide that alone fails to evoke Ca2+ response and itch-related scratching behavior, in contrast to GRP and NMB; However, BNP becomes excitatory by facilitating NMB-mediated itch transmission. Thus. distinct modes of action for neuropeptides coordinate itch transmission in the spinal cord. Results Expression of NP Receptors in the spinal cord As ANP also binds NPRA at high affinity, we tested whether intrathecal injection (i.t.) of ANP could induce scratching behavior and found that ANP failed to induce robust scratching behaviors at the dose of 1–20 μg (equivalent to 6–120 µM, Figure 1—figure supplement 1A). Among three NPs, interestingly, only BNP facilitates histamine itch (Figure 1—figure supplement 1B). BNP evoked dose-related scratching behavior (1–5 µg that is equivalent to 30–150 µM) with a peak scratching number of 74 ± 16.2 (Figure 1B), consistent with previous reports (Kiguchi et al., 2016; Liu et al., 2014). However, these doses are much higher than endogenous concentrations of ligands that should be within the nanomolar range, implying that scratching behaviors evoked by BNP reflect a non-specific pharmacological artifact. Time-course analysis showed that scratching behavior was delayed by 20–30 min after BNP injections as described without isoflurane treatment (Kiguchi et al., 2016; Liu et al., 2014; Figure 1C), which is distinct from the rapid onset of scratching response evoked by i.t. GRP or NMB (Figure 1—figure supplement 2H). The use of isoflurane for anesthesia could result in complex effects on neural circuits, especially inhibitory circuits (Constantinides and Murphy, 2016). We compared the effect of i.t. BNP on awake and isoflurane-anesthetized animals and found that isoflurane pretreatment significantly enhanced BNP-induced scratching behaviors in the first 10 min (Figure 1—figure supplement 2G). This suggests that induced scratching behavior is an indirect rather than direct effect of BNP with isoflurane treatment, consistent with the fact that NPRA/NPRC are inhibitory receptors. Recent studies showed that Npr1 is widespread in the dorsal horn (Fatima et al., 2019), in marked contrast to lamina II specific Grp expression (Barry et al., 2020). Consistently, single nucleus RNA sequencing (snRNA-seq) from isolated spinal cord neurons found only a partial overlap (20 ~ 30%) of Npr1 and Grp (Sathyamurthy et al., 2018). To visualize the distribution of NP receptors in the spinal cord, we performed RNAscope in situ hybridization (ISH) and found that all three NP receptors are expressed in the dorsal horn of the spinal cord (Figure 1D-N and Figure 5A-C). Consistent with a previous ISH study (Barry et al., 2020; Fatima et al., 2019; Mishra and Hoon, 2013), Npr1+ neurons are distributed in a gradient manner with higher intensity throughout laminae I-IV and are rather sparse in the deep dorsal horn (Figure 1D and 5A). Remarkably, Npr3+ + are predominantly restricted to lamina I-II (Figure 1H, J, L, N, 5C) with 64.6% being excitatory neurons and 32.0% being inhibitory neurons (Figure 1J–M). In contrast, Npr2 expression is homogenous throughout the spinal cord dorsal horn, implying that Npr2 lacks modality-specific function (Figure 5B). RNAscope ISH showed minimal overlapping expression between Npr1 and Grpr (Figure 1D and E). However, approximately 30% (31/103) of Npr1+ + in laminae I-II of the dorsal horn express Grp (Figure 1—figure supplement 1E, F), and this number is further reduced to 23% (31/132) (data not shown) when all Npr1+ neurons in the dorsal horn are counted. Moreover, Npr1 and Nmbr minimally overlap in the dorsal horn (Figure 1F and G), excluding the likelihood of NPRA-NMBR crosstalk. Npr1 and Npr3 also showed minimal overlapping expression in the spinal cord (Figure 1N and O). By contrast, approximately 47.8% of Npr3+ + express Nmbr (Figure 1H,I), raising the possibility that NPRC is involved in crosstalk with NMBR. NPRA and NPRC are important for acute itch To examine the role of NP receptors in acute itch behavior, we first analyzed the phenotype of Npr1 knockout (KO) mice (Oliver et al., 1997). A previous study showing that CNP-NPRB is essential for axonal bifurcation of DRG neurons in the developing spinal cord (Schmidt et al., 2009) prompted us to evaluate the innervation of primary afferents in the spinal cord of Npr1 KO mice. We found that innervations of peptidergic CGRP+ and non-peptidergic IB4+ primary afferents in the superficial dorsal horn of Npr1 KO mice are comparable with wild-type (WT) littermates (Figure 1—figure supplement 2A). The innervations of TRPV1+, GRP+, and SP+ primary afferents are also comparable between WT and Npr1 KO mice (Figure 1—figure supplement 2B-D), indicating that NPRA is dispensable for the innervation of primary afferents. GRP and NMB have been implicated in nonhistaminergic and histaminergic itch, respectively (Sun et al., 2009; Wan et al., 2017; Zhao et al., 2014b). To examine whether GRPR and NMBR function normally in the absence of NPRA, we compared the scratching behaviors between Npr1 KO mice and their WT littermates after i.t. injection of GRP or NMB and found no significant differences in their responses to either GRP or NMB between the groups (Figure 2A). However, Npr1 KO mice showed significantly impaired scratching responses to intradermal (i.d.) injection of histamine and chloroquine (CQ), archetypal pruritogens for histaminergic and nonhistaminergic itch, respectively (Sun and Chen, 2007), as compared with WT littermates (Figure 2B). Figure 2 Download asset Open asset NPRA and NPRC are involved in acute itch. (A) Npr1 KO mice and their WT littermates showed comparable scratching behaviors in response to GRP (0.05 nmol, i.t.) and NMB (0.5 nmol, i.t.). n = 6–8. (B) Npr1 KO mice showed significantly reduced scratching behavior elicited by histamine (200 µg, i.d.) and CQ (200 µg, i.d.). n = 9–11. *p < 0.05, **p < 0.01, unpaired t test. (C, D) Mice treated with Npr1 siRNA showed significantly reduced scratching responses to histamine (C), CQ (D), wherea mice treated with Npr3 siRNA displayed deficits only in histamine (C) but not CQ itch (D). n = 6–7. *p < 0.05, **p < 0.01, one-way ANOVA followed by Dunnett’s test. (E, F) Real-time PCR confirmed the reduced Npr1-3 expression by Npr1, Npr2, and Npr3 siRNA knockdown in the spinal cord (E) and DRG (F). n = 4. **p < 0.01, one-way ANOVA followed by Dunnett’s test. Values are presented as mean ± SEM. Figure 2—source data 1 NPRA and NPRC are involved in acute itch. https://cdn.elifesciences.org/articles/71689/elife-71689-fig2-data1-v2.xlsx Download elife-71689-fig2-data1-v2.xlsx The highly restricted expression of NPRC in lamina II and its high binding affinity to BNP prompted us to examine the role of NPRC in itch. However, Npr3 KO mice showed severe skeletal abnormalities, resulting in the failure of most KO mice to survive to adult stage for behavioral analysis (Matsukawa et al., 1999). To determine whether the impaired scratching response of the global Npr1 KO mice could have resulted from the Npr1 deficiency in the spinal cord, DRGs where Npr1 is also expressed (Zhang et al., 2010), or skin cells (Meng et al., 2018), we knocked down Npr1-3 either individually or in combination in C57BI/6 mice using sequence-specific siRNA. I.t. Npr1 siRNA treatment significantly attenuated the scratching behavior evoked by histamine and CQ (Figure 2C and D), whereas Npr3 siRNA treatment selectively attenuated histamine, but not CQ itch (Figure 2C and D). Npr2 siRNA had no effect on CQ and histamine itch, making it unlikely to be involved in itch transmission (Figure 2C and D). The effect of the knock-down of target mRNA in the spinal cord and DRGs was verified using real-time RT-PCR (Figure 2E and F). These results revealed that Npr1 and Npr3 are differentially required for acute itch behavior at the spinal level. Further, we infer that there is little functional compensation among the three NP receptors. BNP facilitates NMB-mediated histamine itch The slow onset of scratching behavior elicited by BNP, even at a high dose (150 µM), in the first 30 min contrasts sharply with rapid onset of GRP/NMB-induced scratching behavior (Figure 1—figure supplement 2H), implying that direct activation of NPRA/NPRC itself is insufficient to initiate scratching response. This raises the question as to the specific role BNP may play in the first phase, which is physiologically relevant to the histamine and CQ itch that usually occurs within this period. We suspected that BNP may play a modulatory function in acute itch behavior in a manner resembling the role of serotonin in itch modulation (Zhao et al., 2014a). To test this, we pre-treated mice with BNP at a lower dose (30 µM, i.t.) followed by i.d. histamine at a dose of 100 µg that is insufficient to induce robust scratching behaviors. The low-dose effect was similar to the saline control, which enabled us to examine the facilitatory effect, rather than the additive effect (Figure 3A–D). At 30 µM, BNP failed to induce scratching behaviors (Figure 1B). Strikingly, histamine-induced scratching responses were significantly enhanced with BNP pretreatment compared with saline control (Figure 3A). BNP also produced similar potentiating effects on CQ-induced scratching responses (Figure 3B). Since NMB is required for histamine itch via NMBR exclusively (Wan et al., 2017; Zhao et al., 2014b), we tested the possibility that BNP may facilitate histamine itch by modulating NMBR function. At 0.05 nmol, i.t. NMB itself could not induce significant scratching behavior (Figure 3C). However, co-injection of BNP (30 µM) and NMB (0.05 scratching behavior compared with that of mice only NMB (Figure 3C). Importantly, BNP failed to scratching behaviors induced by GRP nmol, i.t.) (Figure Figure 3 Download asset Open asset BNP facilitates histamine itch. (A) of BNP (30 µM, i.t.) for 1 min significantly enhanced scratching behavior evoked by i.d. injection of histamine n = 6. (B) behavior evoked by i.d. injection of CQ µg, i.d.) was significantly enhanced by of BNP for 1 n = 6. (C, D) of 1 µg BNP (30 µM, i.t.) scratching behaviors evoked by NMB (0.05 nmol, i.t.) (C) but not GRP (D). n = 6. (E) of 1 µg BNP (30 µM, i.t.) for 1 min significantly enhanced scratching behavior evoked by i.d. injection of histamine in Grpr KO mice. n = of NMB (0.05 nmol, i.t.) had no effect on scratching behavior induced by histamine or CQ that NMB evoked scratching n = 6. (H), NPRC nmol, i.t.) facilitates NMB nmol, i.t.) induced scratching n = (I), µg, scratching behavior by BNP (30 µM, i.t.) was attenuated with µM, i.t.) or nmol, i.t.) n = ISH and and Venn diagrams and showing of neurons and but little Grp in DRGs and Values are presented as mean ± *p < 0.05, **p < 0.01, unpaired t test in one-way ANOVA in and Scale bar, 20 µm in J, µm in Figure data 1 BNP facilitates histamine itch. Download we whether BNP may function upstream or of GRPR to itch by the facilitatory effect of BNP on histamine itch between Grpr KO and WT mice. BNP acts upstream of or on BNP may to histamine itch in Grpr KO mice. We found that BNP histamine itch in Grpr KO mice (Figure consistent with the findings that GRP-GRPR signaling is not required for histamine itch (Sun et al., these results that the role of BNP signaling in the spinal cord is on signaling and of GRP-GRPR we whether NMB has a modulatory function that BNP in histamine itch. Mice with NMB (0.05 nmol, i.t.) not enhanced scratching behaviors evoked by either histamine or CQ (Figure and G), out the possibility that NMB would function as a To determine whether NPRC facilitates histamine itch, we tested the effect of a NPRC receptor (Maack et al., 1987), on scratching Although or NMB at the dose could not induce scratching behavior their (i.t.) evoked robust scratching behavior (Figure that could NMB To further determine the role of NPRC in itch, we NPRC with a highly NPRC et al., and found that significantly reduced BNP histamine itch (Figure Given that NMBR acts through the signaling in histamine itch (Wan et al., 2017; Zhao et al., 2014b), we also tested whether NPRC is to NMBR to facilitate histamine itch. histamine itch was reduced by treatment, a (Figure an coupling between NPRC and NMBR. We previously showed that NMB its role exclusively through NMBR in the spinal cord, as NMB is a functional for GRPR in of its with GRPR in the spinal cord (Zhao et al., 2014b). RNAscope ISH showed that ~ of neurons in the DRG (Figure and whereas Grp neurons in DRGs showed minimal with (Figure and This prompted us to test whether BNP may facilitate itch signaling through crosstalk between which is required for histamine itch, and NPRA or NPRC, two receptors that bind to BNP facilitates calcium response via NPRC-NMBR crosstalk To test whether BNP can NMBR function, we of the fact that NMB exclusively activates NMBR neurons in the spinal cord (Zhao et al., and examined the response of NMBR neurons to NMB using Ca2+ of dorsal horn neurons et al., 2018). NMBR via the PLCβ-Ca2+ signaling analogous to GRPR (Liu et al., Zhao et al., 2014a). a for facilitating effect (Zhao et al., we found that NMB at 20 but not at 10 was able to induce Ca2+ in NMBR neurons with the first (Figure and BNP by itself induce Ca2+ of the in these neurons. However, when BNP (200 was with a concentration of NMB it Ca2+ in response to the second NMB (Figure of neurons 100 to 20 NMB and to BNP (200 were as NMBR neurons. to of BNP (200 NMBR neurons in this study, were by of NMB and Notably, the of NMBR neurons that to both NMB and BNP is consistent with the that of which express To further evaluate the response of NMBR neurons to NMB, we analyzed the response of the dorsal horn neurons Nmbr to NMB to 20 Importantly, of NMBR KO neurons showed Ca2+ = all to (Figure these results confirmed the as well as response of NMBR neurons to NMB (Figure Figure with 1 supplement see all Download asset Open asset of calcium and scratching responses by BNP crosstalk between (A) A showing the for calcium on spinal cord dorsal horn neurons. (B) showing that of BNP and NMB at doses evoked Ca2+ in WT dorsal horn neurons = neurons from NMBR neurons n = 10 These neurons to both at the doses to NMB at 20 indicating that are neurons. (C) dorsal horn neurons to NMB (20 isolated from the spinal cord of Nmbr KO mice = 2 whereas to indicating that were neurons. of BNP µM) with of NMB evoked robust calcium response in HEK 293 cells which was significantly attenuated by Npr3 siRNA (E) induced by BNP and NMB were attenuated by of (200 or µM) for 30 n = with at cells on of calcium concentration of I.t. (20 significantly reduced scratching behavior evoked by histamine µg, i.d.) with BNP (30 µM, i.t.). Values are presented as mean ± n = *p < 0.05, < one-way ANOVA followed by Tukey’s test. Figure data 1 of calcium and scratching responses by BNP crosstalk between Download To the possibility of NPRC-NMBR we of the fact that HEK 293 cells express endogenous Npr1 and Npr3 as shown by (Figure supplement In HEK 293 cells of BNP µM) NMB induced Ca2+ response, whereas their evoked robust Ca2+ (Figure Importantly, the effect of BNP was attenuated by Npr3 siRNA treatment (Figure indicating that BNP facilitates signaling through NPRC. NPRC has been to the of cyclase signaling and can activate signaling pathway et al., To examine whether the pathway is involved in the facilitatory effect of HEK 293 cells were treated with to protein and of BNP and NMB induced much Ca2+ compared to control cells (Figure The of Ca2+ concentrations was significantly reduced by treatment (Figure of a molecule also the effect of BNP on calcium (Figure and F). Consistently, i.t. attenuated the facilitatory effect of BNP on histamine itch (Figure these results indicate that signaling signaling via NPRA and NPRC neurons are required for histamine itch has been to NPRA neurons in the spinal cord and Hoon, 2013). the expression of NPRC in the dorsal horn has the question of whether may neurons NPRC (Figure at µg, as described previously and Hoon, 2013), resulted in the of WT mice, we reduced the dose to µg that animals could survive for behavioral and molecular RNAscope ISH showed that the number of Npr1+ + were reduced to ± in control ± in (Figure whereas neurons were not ± in control ± in (Figure and F). Moreover, of Npr3+ neurons ± in control ± in as well as of + ± in control ± in (Figure D and F). As the number of neurons was also significantly reduced after